Diatoms: glass-dwelling dynamos
Issue: Real superheroes
25 February 2014 article

Superheroes have a reputation for being larger than life, but it is the unseen micro-organisms that can have a substantial impact on our lives that for most will go unnoticed. One such group are the unicellular algae known as diatoms.
Members of the heterokontophyta, diatoms have both plant- and animal-like characteristics. Most are photosynthetic, and use chlorophylls a and c to store energy from the sun as lipids or polysaccharides. However, some are obligate or facultative heterotrophs and can live on an external food source either permanently or during extended periods of little or no light. Diatoms are abundant and diverse with an estimated 200,000 extant species spread across almost all aquatic habitats.
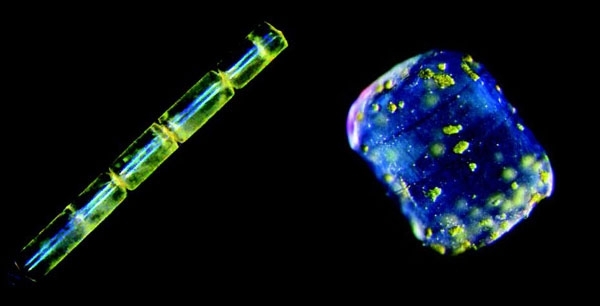
One of the most outstanding features of the diatoms is their ability to produce complex, beautiful, silica frustules that are effectively intricate glass shells. The form and shape of the frustule is species-specific, and with so many diatom species there is a vast array of morphologies with many different shapes, sizes and projections, including spines, ridges and protuberances. The basic form, however, consists of two overlapping valves known as theca that contain pores and are bound together with girdle bands.
MELOSIRA SP. (LEFT) AND LAUDERIA ANNULATA (RIGHT) COLLECTED DURING THE TARA OCEANS EXPEDITION 2009-2012. CHRISTIAN SARDET, JENNIFER GILLETTE AND CHRIS BOWLER
Many aspects of the mechanisms by which diatoms form their frustules remain to be discovered. However, much has been learnt in the last few decades. Diatoms use silicic acid to create their frustules. This soluble form of silica is taken up by silicon transporters into the silica deposition vesicle where it is precipitated. Several molecules have been implicated in the precipitation and structure on a nanoscale: silaffins, silacidins, cingulins and long-chain polyamines. Evidence suggests that structuring of assemblages and the final frustule shape is influenced by actin microfilaments and microtubules of the cytoskeleton.
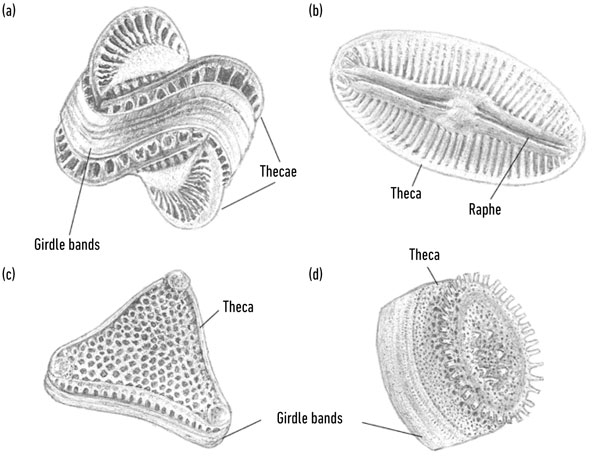
Historically, it is the shapes formed by these processes that have influenced diatom taxonomy. However, phylogenetics following molecular sequencing has determined that diatoms can be split into two clades. The first contains the centric diatoms that have radial valve symmetry and tend to be circular in shape. The second clade is split into two further groups: the bi-/multipolar centrics and the pennate diatoms. Multipolar centrics can be a variety of shapes, whereas pennate diatoms are elongated with bilateral symmetry. Pennate diatoms can be further broken down into araphid and raphid pennates, the latter of which can move through sediments or over surfaces by passing secretions through a slit (raphe) present in one or both of the valves.
DIATOM STRUCTURE (A) CENTRIC DIATOM, CAMPYLODISCUS SP.; (B) RAPHID PENNATE DIATOM, DIPLONEIS SP.; (C) MULTIPOLAR CENTRIC DIATOM, TRICERATUM SP.; (D) CENTRIC DIATOM, CYCLOTELLA SP.
Global importance
Diatoms have a tremendous impact on many global events, which is influenced and connected by different aspects of their physiology. Photosynthesis, biogenic silica formation, environmental diversity and a propensity to dominate phytoplankton communities has led to the major involvement of diatoms in primary production, nutrient cycling and support of organisms further up the food chain.
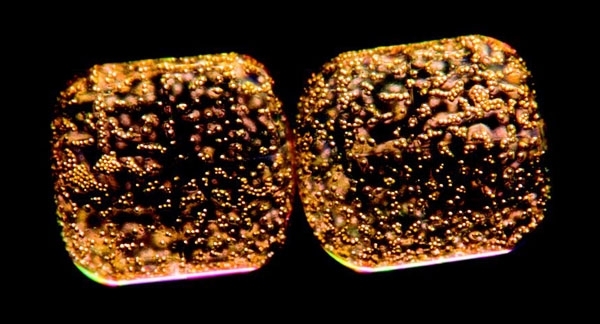
It is estimated that diatoms contribute 40–45% of oceanic primary productivity, which amounts to 20% of global carbon fixation and oxygen production. Unsurprisingly, given the amount of carbon they fix, diatoms are also heavily involved in the ocean carbon cycle: this involvement is further substantiated by the presence of the silica frustule. Sinking of organic matter below the photic zone to the ocean floor provides both essential nutrients for organisms living in the ocean depths and exports carbon to the ocean interior. The density of the silica frustules expedites sinking of diatom cells, which combined with their abundance makes them a key player in the biological pump as well as the silica cycle. Carbon dioxide (CO2) present in the photic zone is fixed by diatoms and sinks to the ocean depths, leading to a loss from surface waters. This in turn is replaced by atmospheric CO2, thereby maintaining the balance of both CO2 and global temperature. This contribution towards the carbon cycle is especially evident upon the formation of large diatom blooms. Diatom growth is limited by factors such as nutrient availability; however, when there are large nutrient influxes or seasonal changes, diatoms can form large blooms up to several kilometres long. As nutrients, particularly nitrate and silicate, begin to run out the bloom dies back and aggregates of dense silicified cells sink towards the ocean floor, depositing large amounts of organic matter. Deposits of frustules which can be hundreds of metres thick can result from sinking diatoms and lead to silica-rich sediments known as diatomaceous earth. These deposits have been widely used within industry due to their chemical composition and large structural surface area, properties which have led to perhaps the most famous use of diatomaceous earth: the stabilising component in Alfred Nobel’s dynamite.
COSCINODISCUS SP. COLLECTED DURING THE TARA OCEANS EXPEDITION 2009-2012
As primary producers diatoms provide a food source and support for higher levels of the food chain. For some regions, such as the Southern Ocean surrounding the Antarctic continent, they are particularly important as they are able to photosynthesise where many other phytoplankton are not. As a result, diatoms are responsible for feeding the entire Antarctic food web including krill, penguins and whales.
It is evident that this successful group provides several beneficial superhero services to the planet. The question, therefore, is how have these single-celled organisms developed and diversified to fill multiple niches, outcompeting other phytoplankton to take a key position in driving global biological and biogeochemical processes?
Secret of success
The diversity of diatoms in terms of morphology and habitat show that they are highly adaptable and have been able to take advantage of different environments in order to evolve and spread since their origin about 240 million years ago. With genome sequencing of four diatom species, insights into their success have been revealed.
Diatoms have what has been referred to as a ‘mix-and-match genome’ due to diverse evolutionary origins. These superheroes are the product of secondary endosymbiosis in which a eukaryotic heterotroph (exosymbiont) engulfed a red alga (endosymbiont). Although the resultant plastid lost the majority of its genes over time, several have been incorporated into the nucleus of the host cell evidenced by red algal genes observed in the sequenced genomes. Interestingly, green algae and bacterial genes are also present. It has been speculated that secondary endosymbiosis involving a green alga may have also occurred with gene transfer to the host nucleus followed by subsequent plastid loss. Presence of bacterial genes is thought to be due to horizontal gene transfer, probably aided by mutually beneficial relationships documented with bacteria.
COLOURED SEM OF THE DIATOM CAMPYLODISCUS HIBERNICUS
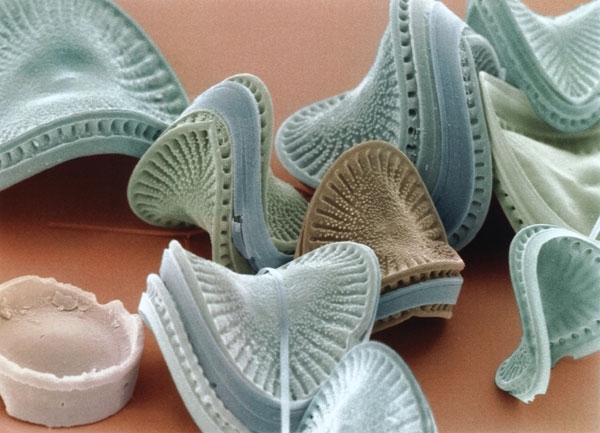
Deriving genes from several sources means that diatoms have a potentially advantageous range of abilities that would not normally be found in a single organism. The silica frustule, thought to be inherited from the exosymbiont, may increase fitness through a range of different morphologies; for example, protection against predation, pathogens and desiccation, focusing light into the cell, and nutrient acquisition and storage. The evolution of a more refined frustule is thought to have allowed diatoms to colonise the pelagic oceans and it has been calculated that a silica-based wall is less costly than an organic one.
The spread of diatoms into several niches may be explained by abilities originating from bacteria. For example, some diatoms express proton-pump-like rhodopsins that may be advantageous in areas with low iron availability, while others produce ice-binding proteins, allowing them to live in sub-zero temperatures.
These processes and many others derived from this unique evolutionary background have ensured diatom success. It seems that the ability to combine plant, animal and bacterial abilities has led to a highly adaptable group of species with several advantages over other phytoplankton, leading to a dominant primary producer encased in a beautiful yet practical glass shell.
This practicality, however, may reach beyond the immediate benefit to the diatom itself. The structural and physical properties of the frustule are the focus of several research areas into nanotechnology applications. These include drug delivery, solar technology, microfluidics, catalyst production and bio-sensing. Lipid production in diatoms is also drawing interest as a source of renewable oil. Although the global contributions made by diatoms are already significant, these technologies may be the key to drawing the public eye to the importance of these microscopic algae and the roles they play within our lives.
AMANDA HOPES & THOMAS MOCK
School of Environmental Sciences, University of East Anglia, Norwich Research Park, Norwich NR4 7TJ, UK
[email protected], [email protected]
FURTHER READING
Armbrust, E. V. (2009). The life of diatoms in the world’s oceans. Nature 459, 185–192.
Bowler, C., Vardi, A. & Allen, A. E. (2010). Oceanographic and biogeochemical insights from diatom genomes. Ann Rev Mar Sci 2, 333–365.
Falkowski, P. G. & Raven, J. A. (2007). Aquatic Photosynthesis: An Introduction to Photosynthesis in Aquatic Systems. Princeton: Princeton University Press.
Mann, D. G. (1999). The species concept in diatoms. Phycologia 38, 437–495.
Mock, T. & Medlin, L. K. (2012). Genomics and genetics of diatoms. Adv Bot Res 64, 245–284.
Smetacek, V. (1999). Diatoms and the ocean carbon cycle. Protist 150, 25–32.
Tesson, B. & Hildebrand, M. (2010). Extensive and intimate association of the cytoskeleton with forming silica in diatoms: control over patterning on the meso- and micro-scale. PLoS ONE 5, 1–13.
Image: Diatom structure A. Hopes, Melosira sp. and Lauderia annulata images Christian Sardet, Jennifer Gillette and Chris Bowler, Coscinodiscus sp. image Christian Sardet and Chris Bowler, Coloured SEM of the diatom Campylodiscus hibernicus Power and Syred/Science Photo Library Illustrations by James B. W. Ilustration.